| 羽化・ライフサイクル | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
羽 化 鹿児島では四月初旬、大分以北以東では四月中旬から下旬に羽化を始め、天候により20日前後のずれを生じます。 羽化開始から羽化終了までは約一ヶ月程度ですが、平均して羽化開始から約4日目に羽化のピークがあり、このピークは約8日間続きます。 但し、それは好天が続いた場合で、低温や雨が続くとこのピークはなくフラットな状態が続きます。 ピーク後、急激に羽化数は減少しますが、それでも、わずかずつだらだらと見られます。 羽化時刻はAM:06時頃を中心に2時間程度の幅があるものと考えられます。 羽化後の初飛行は10時から12時の間がピークになります。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
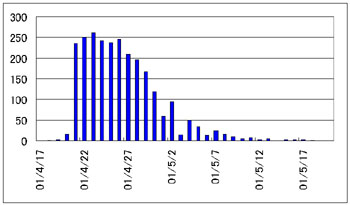 羽化個体数の推移 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
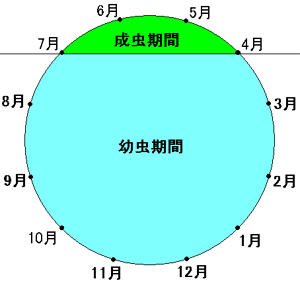 ライフサイクル |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 繁殖行動 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ベッコウトンボ羽化期の野依新池(大分県中津市)  右手のススキの立ち枯れが見える土手の向こうはベッコウトンボの生息池 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
前繁殖期 ベッコウトンボは羽化すると数時間羽化水域にとどまり、やがて水際のへ移動します。午前12時頃を過ぎるとさらに水辺から離れ、ススキやチガヤなどが立ち枯れる草地へと移動します。 ベッコウトンボの燈色に黒の体色は枯れたチガヤの中に入ると見事にまぎれます。 羽化水域から離れた草地へ移動する習性はオス・メスに等しく認められ、オスの場合は性的に成熟するまでの期間、メスの場合は生殖行動をしない時は常時この草地で過ごします。 繁殖行動に参加しないこの期間のことを前繁殖期といっています。 ベッコウトンボが成熟するまでに要する期間は9日から11日で、オスメスの差はないと考えられます。 繁殖期 性成熟した(オスは精嚢が成熟し、メスは卵巣が成熟した状態)オスは羽化水域に戻りヒメガマ等の枯れ茎に静止して縄張りを形成します。この時の静止姿勢は水平に近く、4脚で止まります。 しかし、ベッコウトンボの縄張りに対する執着は強くなく、この性向が繁殖地における高密度の生息を可能にしています。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 幼虫-1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
幼虫の飼育については2004年の飼育結果を基に、1993年の近藤祥子・青木典司氏のレポート及び発生は渡辺庸子氏のレポートを参考にして紹介します。 孵化は前幼虫の形でおこなわれ、体が卵から出るとすぐに体が卵殻にひっかかっいる状態で前幼虫の背中が割れて1齢幼虫が出てきます。 終齢幼虫になるまでには11回脱皮し12齢で終齢幼虫になります。各齢期の体長と体幅を以下に示します。餌は飼育時に与えたもので、自然条件下の時はこのようではありません。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
自然状態では10月の終わり頃にはほぼ終齢幼虫に達しているようですが、遅くとも12月には全て終齢に達します。 室内での飼育下では8月25日に終齢に達した個体があり、9月終わりには85パーセントが終齢に達しました。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
  左1齢幼虫 右4齢幼虫 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
幼虫の餌 1 6齢以降はセスジユスリカを与えました。多数のヤゴを養殖する時にはユスリカも同時に養殖すると餌を与えるのに便利です。飼育容器の中には水草をいれてもい餌が切れても餌となる生物が発生しているような状況を作っておくと良いでしょう。こうすることで一種だけの餌を食べる事による摂取物の偏りも防げます。 幼虫の餌 2 写真はフサカ幼虫の頭部と胸部です。 顕微鏡下ではじめてこれを見たときは宇宙人かと思いました。胸のブラジャーのように見えるのは浮き袋です。生きている時はこのようにしわだらけではありません。アルコールに浸したためにこのようになってしまいました。 このフサカの幼虫はワムシを食べる小型のミジンコ類を捕食し、池の中の小型ミジンコ類の密度を低下させます。(1998年 花里)その結果フサカが多い池ではワムシの密度が高くなるのです。 ベッコウトンボが安定して生息している中津市の野依新池では水深10cm~30cmの浅い水域に高密度でフサカの幼虫が生息しています。 弱齢幼虫を生息地の泥や水底の植物と一緒に顕微鏡下で観察した時、小型のワムシを好んで食べている様子が観察できました。 フサカはカやユスリカの仲間で終齢幼虫の大きさは10mm程度、無色透明で水中では横になって浮き、近づいてくるミジンコ類を捕食します。フサカ自身もベッコウトンボ幼虫の餌になっているのではないかと考えています。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
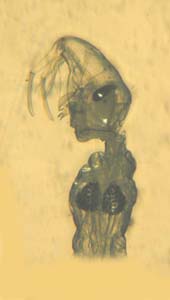 フサカの幼虫 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 フサカ幼虫はこのような姿をしています。見事な透明です。 この写真はホームページ「konekoの森」に掲載されているフサカ幼虫です。 http://www.hpmix.com/home/tenaga/konekonomori2/index.htm |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 フサカ幼虫の尾部です。扇状に広がっているのは遊泳毛。 この写真はホームページ「konekoの森」に掲載されているフサカ幼虫です。 http://www.hpmix.com/home/tenaga/konekonomori2/index.htm |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 フサカ幼虫の弱齢個体です。胴体部分が短いですね。 この写真はホームページ「konekoの森」に掲載されているフサカ幼虫です。 http://www.hpmix.com/home/tenaga/konekonomori2/index.htm |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 幼虫ー2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 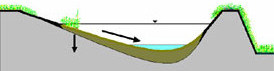 水が引いた底が水平な池 傾斜がついた池 水が引いた底が水平な池 傾斜がついた池 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
減水時の行動 ベッコウトンボの生息環境は大型抽水植物が一面に広く生い茂った池沼です。大型抽水植物は一定の水深までしか生育できません。従って、その植物が水面を広く覆っているということは水深の差は小さく、水底は水平に近いということです。 池の水が干上がると、池に棲む生物は水際線に沿って後退します。 即ち、水平移動です。シオカラトンボやギンヤンマ等多くのトンボの幼虫はこのような行動を取ります。しかし、ベッコウトンボやヨツボシトンボ等は水がなくなるとそのまま泥に潜ります。垂直移動です。(青木,) 水平な水底を住処とするトンボにとっては水が干上がった時に横に動いても結果は同じなのです。 わずかに残った水を求めて動き回るエネルギーのロス。捕食者に捕食される危険性の高さ。こう考えればその場で泥に潜るという選択は最善の選択と言えます。 しかし、水底が傾斜している場合は最悪の選択となる事はいうまでもありません。 種間競争 ベッコウトンボと同属のヨツボシトンボとの間の種間競争の問題がよく論じられます。ヨツボシトンボとベッコウトンボの幼虫を一緒に飼育するとベッコウトンボはヨツボシトンボに食べられてしまうという報告があります。報告者も述べているようにそれは飼育槽という特殊な環境下でのことです。 実際の生息環境である池の水底はヒメガマなどの抽水植物の枯れ茎が幾重にも積み重なっています。 水槽が何もないグラウンドだとすれば実際の生息環境は高層ビルの中のようなものです。実際にはヨツボシトンボによる捕食圧は低い思っています。 共食い ベッコウトンボの幼虫には共食いは見られないという意見もありましたが、幼虫の生息密度が高くなれば共食いは頻繁に見られます。 但し、ベッコウトンボの幼虫は行動が不活発であまり動きません。従って、他個体と出会う確率は活発に動き回る種に比べると低く、共食いする確率も小さくなります。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 孵化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
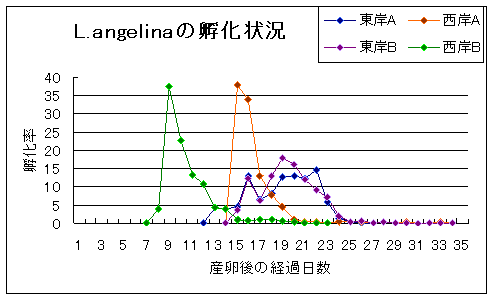 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 上に示したグラフは孵化の記録です。卵数は何れも各1000~1200個。採集は大分市6号埋立地にある池で採集しました。 池はほぼ四角形で、池の西岸沿いの草地で2個体のメスを、東岸沿いの草地で2個体のメスをそれぞれ採集しました。 西岸Bは28℃に保った水温での結果です。その他の3サンプルは室温に放置しました。 その結果、東岸グループと西岸グループでは孵化のパターンに際立った違いが見られました。 この事は一つの池の中であっても異なった遺伝的特質を持つ複数のグループがエリアごとに存在している可能性を示唆しています。 ただし、このデータは数の面からも精度の面からも不満足なものであり、予備調査的な性格のものであることをお断りしておきます。 はたして、自然環境下ではどのような違いとなって現れるのか、これこそが正に知りたいところです。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 内部生殖器 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
ベッコウトンボ♀腹部断面(輪切り) 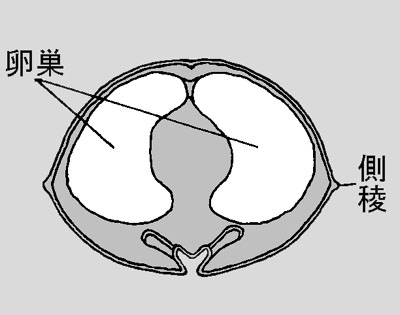 メスの腹部は大部分が卵巣で占められています。中央の空隙に消化器系器官が通っています。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
卵巣と輸卵管の位置(平面・縦断) 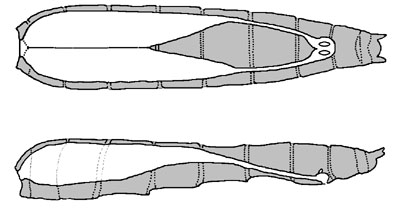 性成熟すると卵巣は図のように大きく膨れ、輸卵管も卵で膨れる。輸卵管の末端には交尾嚢と貯精嚢があります。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
ベッコウトンボの卵巣  卵巣を取り出したものです。取り出す際に卵巣の一部を傷つけてしまい、成熟卵や卵巣小管が外に飛び出してしまいました。 卵巣小管 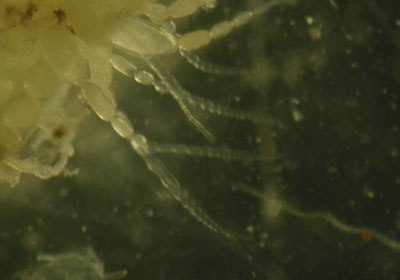 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 卵巣内には数百の卵巣小管があり、一本の卵巣小管には約30個の卵が連なっています。 卵は根元から順に成熟し、成熟した卵は外に飛び出す。 老熟したメスでも多数の未成熟卵を持っているので産卵に利用するのは20~30%ではないだろうかと考えています。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
産 卵 メスは原則として交尾後、直ちに打水産卵します。しかし、他のオスの干渉が激しかったりすると産卵半ばで繁殖水域から離脱します。 その後、輸卵管が膨満して産卵意欲が高まってくると再度繁殖水域に侵入し、この時オスに見つからなければ単独で産卵します。 また、一度交尾を経験したメスは精子を貯精嚢に保存しているので、それ以後は交尾なしでも産卵できますが、産卵のたびに他のオスと交尾した方が多様な子孫を残す事が出来ます。 6月にはいると繁殖水域の個体密度は低下します。このような時、老熟したメスが単独で産卵しているのをよく見かけます。 寿命 寿命は約30日程度ではないかとの見方が一般的ですが、マーキングによる再捕獲記録では50日・60日の記録もあり意外に長寿の個体があるのかもしれません。 マーキングにより平均寿命を調べる場合、マーキング行為が個体へ与えるダメージによって短命になる可能性も考えられることから調査方法を検討する必要があります。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
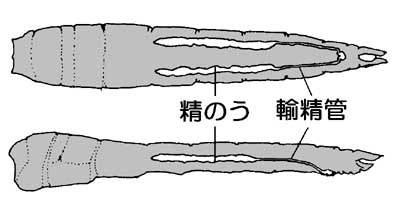
ベッコウトンボ精嚢 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| トンボ科のものはほぼこのような形です。 精子の形は両端がとがったステッキ状をしています。 輸精管の出口は観音開きのドアになっています。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 分散 - 移動 と 移出 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ベッコウトンボは生息地にとどまる性質が大変強いことが知られていて、この性質がベッコウトンボの大きな特徴となっています。 一方では分散する性質についても幾つかの報告があります。この項ではベッコウトンボの分散する性質についてまとめてみます。 分散には二つのパターンがあります。 一つは発生した水域から離れて草地へ移動するものでリターンを前提にした移動です。 オスでは前繁殖期に見られ、メスでは前繁殖期・繁殖期を通じて見られます。この時の移動距離は周辺に良好な草地がどれだけあるか(個体群の規模に比して)に影響されますが、500mを超えることはないようです。 もう一つはリターンを伴わない新しい生息地を求めての移動で、これを前述の移動と区別するために移出と呼びます。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
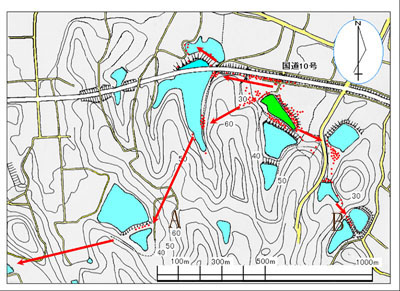
上の地図は大分県中津市野依の新池で羽化した個体群の移動の状況を表しています。 この地域にはさまざまなタイプの池がありますが、ベッコウトンボが生息しているのは緑色に塗った野依新池だけです。 ただし、生息は確認されていませんが移出個体と思われるオスがナワバリ行動をしている池は他に2ヶ所あります。 図のA地点やB地点へ移動した個体は更に以遠の池へ移動し移出個体となります。 移出がなぜ発生するのか考えて見ましょう。 一つは遺伝的な性質によるという考え方があります。 もう一つは草地をたどって移動した結果、生息地から離れ過ぎてしまったという考え方です。 羽化直後の個体が処女飛行でいきなり数十メートルの上空へ舞い上がり、上空の強風に飛ばされて一瞬のうちに見えなくなるという例も幾つか目撃しています。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 個体群の構造 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
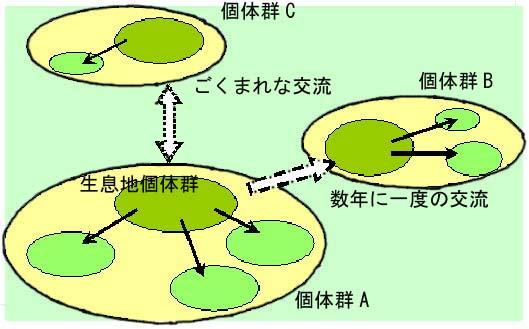
ベッコウトンボの理想的な生息地に見られる個体群構造の模式図です。おそらく、1950年代以前のベッコウトンボ生息地は孤立したポイントとして存在するのではなく、図のように面として、あるいは、ネットワークとして存在したのだろうと考えられます。 有力な生息地個体群は生息密度が高く、生息地から離れる個体も多いと考えられます。 そのため、周辺にある抽水植物が生育する池にもベッコウトンボが見られます。 この場合、有力な発生地から約2km以内の場合は毎年必ず交流があります。 それ以上距離が離れた場合は供給する側の生息密度や規模さらに風向や地形により状況は変化します。 状況によっては30kmや40kmの移動は決して珍しいものではありません。但し、移動した結果、生息に適した池に到達できる可能性は奇跡的なものになるのでしょう。 他の個体群との間で交流が望めない孤立した個体群においては新たな遺伝子の供給ができず、遺伝的に偏った個体群になる恐れがあり、このことが、ひいては生息地の消滅につながると考えられています。 ところが、孤立した個体群であっても平衡が保たれた安定した状態にある個体群も存在する事が判って来ました。 鹿児島県祁答院町の藺牟田池は火口湖の中にあり周辺に生息地はありませんが、個体群は安定しています。 この個体群は多様な遺伝子型を内包した個体群ではないかと推察されます。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 成熟と黒化 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| その1 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
トンボ目に属するトンボは多くの種で成熟すると体色が変化します。しかし、変化の度合いはオスメスや種によって様々に違います。 ベッコウトンボにおいてもこの体色変化は顕著で、変化は黒化という形で現れます。 黒化は前後翅の翅脈、胸部、腹部、複眼、前額等ほぼ全ての部位で見られ、羽化後の経過時間に比例して黒化が進行します。 10年程前までベッコウトンボは「前縁脈の黒化 = 成熟」すなわち、前縁脈の黒化が完了した時点で成熟するとみなされていました。 しかし、どうも少し違うらしいことに気が付き黒化と生殖器官の成熟度、及び捕獲時の行動を照らし合わせた結果、前縁脈の黒化は成熟後しばらくして始まり、翅脈の黒化が終了するのは老熟の域に達してからという事が判ってきました。 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 成熟と黒化 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| その2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||